Download Enzymatic Removal of Initiator RNA in Mammalian Okazaki Fragment Processing and more Study notes Biochemistry in PDF only on Docsity!
Proc. Natl. Acad. Sci. USA Vol. 91, pp. 9803-9807, October 1994 Biochemistry
Enzymatic completion of mammalian lagging-strand
DNA replication
JOHN J. TuRCHI*, LIN HUANGt, RICHARD S. MURANTEt, YONG KIMt, AND ROBERT A. BAMBARAtt
*Department of Biochemistry and Molecular Biology, Wright State University, Dayton, OH 45435; and tDepartment of Biochemistry, University of Rochester, Rochester, NY 14642
Communicated by Fred (^) Sherman, June (^) 8, 1994 (received for review (^) April 28, 1994)
ABSTRACT Using purified proteins from calf and a syn- thetic substrate, we have reconstituted the (^) enzymatic reactions required for mammalian Okazaki fragment processing in vitro. The required reactions are removal of initiator RNA, synthesis fromanupstre fragment to generate a (^) nick, and then (^) ligation.
With our substrate, RNase H type I (RNase HI) makes a single
cut in the initiator RNA, one nucleotide 5' of the RNA-DNA
junction. The double^ strand^ specific 5'^ to^ 3'^ exonuclease^ removes
the remaining monoribonuceotide. After (^) dissociation ofcleaved RNA, synthesis by DNA polymerase generates a nick, which is then sealed by DNA igase I. The unique specificities of the two nucleases for primers with initiator RNA (^) strongly suggest that they perform the (^) same reactions in vivo.
Considerable progress has been made in the last decade toward identifying and (^) characterizing the (^) proteins involved in mammalian DNA replication (reviewed in ref. 1). A substantial proportion ofthis information is derived from reconstitution of simian virus 40 DNA (^) replication in vitro. The results support a two-polymerase system for replication ofnuclear (^) DNA, with
polymerase 8 and polymerase a/primase responsible for lead-
ing- and lagging-strand synthesis (2, 3). The recent demon-
stration that these enzymes, plus DNA (^) polymerase e, are all required for viability in yeast (^) (4-6) has (^) prompted several proposed roles for the latter (^) enzyme in DNA (^) replication
(7-10). While the viral model has clarified priming and elon-
gation steps in the replication fork, the exact steps involved in
completion of lagging-strand synthesis are not yet clear.
Simian virus 40 reconstitution reactions (^) (11-13) indicated roles
for the mammalian 5'-to-3' exonuclease, a nuclear DNA
polymerase, DNA^ ligase, and^ possibly RNase H^ type I (RNase
HI) injoining of nascent segments of the lagging strand. DNA
ligase I, rather than^ III, was^ shown to be required (12, 13). The
specific function of RNase HI and the cleavage mechanism involved remained (^) to be (^) determined. In Escherichia (^) coli, initiator RNA (^) of Okazaki fragments is thought to be removed by the 5'-to-3' exonuclease of DNA polymerase I, although a role for RNase H cannot be totally excluded (14). Double strand-specific 5'-to-3' exonucleases, which appear to be functional homologs of each other, have been isolated from human cells, mice, and calf (11-13, 15, 16). The calf nuclease displayed an endonuclease function that cleaves the unannealed 5' tail ofa primer on a template, in the presence of a (^) directly adjacent upstream primer (17). This very specific function is (^) also exhibited by E. coli DNA polymerase 1 (18), which (^) strongly suggests that the bacterial and mammalian enzymes are also functional (^) homologs. Mammalian RNase H enzymes have been identified and characterized in detail. RNase H enzymes can be placed into one of two classes (I and II) based on subunit structure, peptide molecular (^) mass, and the ability to use manganese as a cofactor (^) (19, 20). They have been postulated to be involved
in DNA (^) replication and (^) transcription (21), but no specific roles have been established for either class. We had shown (22) that the calf 5'-to-3' exonuclease could work with calf DNA polymerases a, 8, or e and DNA (^) ligase I to (^) join two DNA (^) primers annealed to a template with a four-base gap. In addition, we have demonstrated that 5'- to-3' exonuclease activity is stimulated by synthesis from an upstream primer and that the stimulation is the result of the formation of a nick between the two primers, which is a preferred substrate for the exonuclease (17). These results suggest that^ a^ nick^ translation process occurs prior to the joining reaction catalyzed by DNA ligase I. Goulian et al. (15) previously showed that a combination of RNase HI and 5'-to-3' exonuclease can remove initiator RNA from (^) primer- templates in vitro. Here we define the specific reactions by which these two nucleases can remove initiator RNA in a model Okazaki fragment system before polymerization can make the substrate for (^) ligation.
MATERIALS AND METHODS
Protein Purification. DNA polymerase E, DNA ligase I, and calf 5'-to-3' exonuclease were purified from calf thymus as described (22). Calf (^) thymus RNase HI (^) was purified by adapting the^ procedure reported by Eder and Walder (23) for use with calf thymus. Frozen fetal thymus (400 g) was homogenized in 1.2 liter of buffer A (10 mM Tris, pH 8.4/0. mM (^) EDTA/0.5 mM (^) EGTA/1 mM dithiothreitol) containing
50 mM KCl. The homogenate was centrifuged at low speed
(9000 x g) for 30 min, and then the supernatant was centri-
fuged at 100,000 x (^) g for 60 min. The (^) supernatant from high-speed centrifugation was filtered through four layers of cheese cloth and loaded onto a DEAE-Sepharose column (500 ml) equilibrated with buffer A (^) containing 50 mM KCl. The (^) protein that (^) flowed through the column was applied directly to a CM-Sepharose column (500 ml) equilibrated with buffer A containing 50 mM KCl. The column was (^) developed with a 2.5-liter (^) gradient from 50 to 400 mM KCl. Fractions containing RNase H activity were pooled and precipitated
with 60%o ammonium sulfate. The protein pellet was collected
by centrifugation and dissolved in buffer A (^) containing 1 M ammonium sulfate. The (^) protein was loaded on a 40-ml phenyl-Sepharose column and (^) developed with a decreasing salt gradient from 1 M to 0.4 M ammonium (^) sulfate in buffer A. Active fractions were pooled and (^) dialyzed versus 10
volumes of buffer A containing 10%6 (vol/vol) glycerol and
100 mM KCl. The RNase H activity flowed through both Mono Q and Mono S columns. The flow-through protein from these columns was then applied to a 1-ml heparin-Sepharose
column equilibrated in buffer A containing 10%o glycerol and
100 mM KCl. The (^) activity was (^) eluted with a 15-ml gradient
Abbreviation: RNase HI, RNase H type I.
*To whom reprint requests should be addressed at: University of
Rochester Medical (^) Center, Box (^) 607, 601 Elmwood Avenue, Roch- ester, NY 14642.
9803
The publication costs of this article were defrayed in (^) part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. (^) §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 91 (1994)
from 100 to 600 mM KC1 in buffer A containing 10%1 glycerol. Active fractions were pooled, diluted to a conductivity equal to buffer A containing 100 mM KC1 and applied to a 1-ml blue-Sepharose column equilibrated in the same buffer. RNase H activity was eluted from the column with buffer A containing 500 mM KCl and 2 M MgCl2. Active fractions were dialyzed versus buffer A containing 50%o glycerol and frozen at -70TC. The final pool of RNase HI had a specific
activity of 30,000 units/mg as measured on poly([3H]rA).
oligo(dT) according to Eder and Walder (^) (23). Lagging-Strand Substrate Construction. The (^) lagging-strand substrate was (^) prepared by PCR amplification of a 130-bp region of the plasmid (^) pBS(+) (Stratagene) encompassing the multicloning site and^ T3 RNA^ polymerase promoter. The oligonucleotides used for PCR of the pBS(+) plasmid were DNA primer 1 (5'-ATGTTAGGCGCCGGGCGAATTC- GAGCTCGG-3') and DNA primer 2 (5'-AACAGCTATGAC- CATGATTA-3'). The oligonucleotides were synthesized on an Applied Biosystems 360 oligonucleotide synthesizer and purified by 15% polyacrylamide/7 M urea gel electrophore-
sis. PCR reactions were performed for 30 cycles, and the
130-bp product was purified by (^) electrophoresis through 4% NuSieve GTG (^) agarose (Seakem). The (^) purified double-
stranded 130-mer was then subjected to single-primer ampli-
fication using only primer 1. The single-strand 130-mer tem-
plate DNA product was purified by the same method. In vitro transcription from the T3 RNA polymerase promoter was
performed after HindIII or Pst I digestion to yield RNA
primers of 13 and 21 nucleotides, respectively. RNA primers
were purified by 15% polyacrylamide/7 M urea gel electro-
phoresis. RNA primers were hybridized to the (^) single-strand 130-mer template and were extended (^) by using Sequenase' (version 2.0) and dNTPs. After (^) extension, the (^) substrate was digested with^ Nar^ I, producing a^ 3' recessed terminus, which
was labeled by extension with dCTP and [a-32P]dGTP (
Ci/mmol; 1 Ci =^37 GBq). A 20-nucleotide-long DNA primer
was hybridized to the 130-mer DNA so that its 3' terminus was 29 nucleotides upstream of the RNA-DNA fragment. This substrate is depicted in Fig. 1.
Enzymatic Reactions. Lagging-strand reactions were per-
formed in 20 mM Hepes, pH 7.0/1 mM dithiothreitol/10%o
glycerol/1 mM ATP/10 mM MgCl2/0.1 mM dNTPs in a
volume of 20 ,ul. Reaction mixtures were incubated at (^) 37°C
A 20 DNA (^) 'a
29 13 1*^58 pD RNA DNA
I
orp^ _ GG (^) CG CC GC DNA (^) Nar I (^130 4) Polymerase + dCTP+ dGTP
1 60 -)'I
p RNA^ DNA^ * Pp _ GGCG CCGC
G CCTAACAT C GGATTGTA I*
DNA 122 B 13merRNA 21 mer RNA (^5) -GGGAACAAAAGCU'UGCAUGCC'GCAGGTCGACTCTAGAGG
ATCCCCGGGTACCCAGCTCGAATTCGCCCGGCGCCTAACAT-
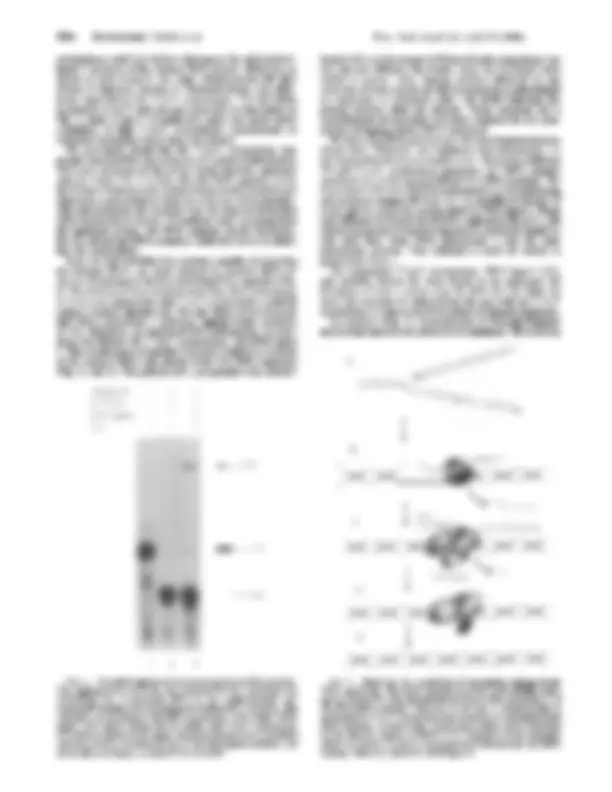
FIG. 1. Model lagging-strand substrate. The (^) lagging strand sub- strate was constructed as described in text. (A) The lengths of segments in^ this and subsequent figures are given in nucleotides. The staggered cut^ by Nar I^ is^ shown. DNA synthesis adds two nucleo- tides, with the label^ on^ the 3'^ nucleotide. The final substrate has a 13-nucleotide RNA attached (^) covalently to a 60-nucleotide DNA. (B) The sequence of the RNA-initiated DNA (^) segment is shown. The covalent RNA-DNA (^) junctions for the 13-nucleotide RNA used in most experiments and the 21-nucleotide RNA used in the (^) experiment in Fig. 4 are indicated.
2 3. C 5
inM- 73 III, b
- 61n .e:*w 63
FIG. 2. An activity with the ability to (^) degrade initiator RNA primers can be purified away from calf 5'-to-3' exonuclease. The 3' labeled lagging-strand substrate was incubated in (^) 20-pl reaction mixtures with various preparations of 5'-to-3' exonuclease purified from calf thymus. Reaction products were separated by 10% poly- acrylamide/7 M urea DNA sequencing gels, dried, and (^) detected by autoradiography. In each lane the amount and specific activity (^) ofthe 5'-to-3' exonuclease preparations in units and (^) units/mg ofprotein are as follows: no enzyme (^) (lane 1), 0.8 and (^) 29,000 (lane 2), 0.4 and 20,
(lane 3), 0.8 and 85,000 (lane 4), and 0.8 and 180,000 (lane 5). The
lengths of the RNA-initiated DNA strands are given at the left. The wide and narrow black lines represent RNA and DNA respectively, in this and subsequent figures. Sizes are shown in nucleotides.
for the time indicated in the figure legends, and reactions were stopped by the addition of 90%o (^) formamide/10 mM EDTA. Reaction products were heated at 95°C for 5 min and separated by denaturing (^) polyacrylamide gel electrophoresis. Products were visualized by autoradiography by using Du- Pont Cronex Lightning Plus intensifying screens at -70°C.
RESULTS AND DISCUSSION
Our previous results have shown that calf 5'-to-3' exonu- clease can (^) degrade a downstream DNA primer and, in conjunction with polymerization by DNA polymerase a, (^) 8, or E, could generate a product that is capable of beingjoined by DNA ligase 1 (22). Therefore, we assessed the ability of the exonuclease to (^) degrade an (^) initiator RNA primer using the substrate shown in Fig. 1. This substrate contains a triphos- phorylated 5' terminus similar to initiator RNA primers synthesized by DNA polymerase (^) a/primase (24). The 3' end ofthe (^) RNA-initiated DNA strand was labeled, so that the fate of the 3' portion of the segment could be evaluated. Fig. 2
RNase HI
ONOW.-W.-4w to _^ -^6
(^2 3 4) >
FIG. 3. Purified calf thymus RNase HI (^) degradation of initiator RNA primers. Calf thymus RNase HI was (^) purified to a (^) specific activity of 30,000 units/mg of protein as described in text. The reactions were incubated for 15 min at 37°C and processed as described in Fig. 2. Lanes: 1, no enzyme; 2, 0.08 units; 3, 0.15 units; 4, 0.3^ units; 5, 0.6 units.^ Sizes are shown in nucleotides.
(^9804) Biochemistry: Turchi et al.
Proc. Natl. Acad. Sci. USA 91 (1994)
exonuclease could not initiate cleavage at the triphosphory- lated 5' terminus of the initiator RNA primer. However, as shown in lanes 4 and 8, the single ribonucleotide left after RNase H digestion, having a 5' (^) monophosphate, was effec- tively removed by the 5'-to-3' exonuclease. The first DNA nucleotide at the 5' end was also removed, as seen faintly in Fig. 5, lanes 4 and 8. Considerably more was made under conditions of high 5'-to-3' exonuclease concentration or extended incubation times (data not shown). We previously found that the 5'-to-3' exonuclease was greatly stimulated by the presence of a primer (^) hybridized to DNA just upstream of the primer being digested, separated only by a^ nick (17). It is likely that the RNA segment cut by the RNase HI temporarily remains bound to the template and stimulates exonucleolytic removal of the last ribonucleotide. This interpretation also explains why the removal of the first DNA nucleotide is slower. If synthesis were occurring from the upstream primer, the RNA segment should dissociate, but the advancing DNA terminus could also serve to stimu- late the exonuclease. Since we had identified the (^) enzymes capable of (^) removing the (^) initiator RNA, we (^) could attempt to combine RNA re- moval with joining of the two remaining DNA^ segments (Fig. 6). We previously have demonstrated that DNA^ polymerase a, 8, or e, in conjunction with 5'-to-3' exonuclease, could fill a gap to support ligation (22). We and others have proposed that DNA polymerase e performs lagging-strand synthesis (8-10). Therefore, we employed DNA polymerase E in reac-
tions with RNase HI, 5'-to-3' exonuclease, and DNA ligase
I. This combination of purified enzymes resulted in removal of the initiator RNA and joining of the two DNA segments (Fig. 6, lane 3). The polymerase e preparation was contam-
RNase Hli 51 -^ E X .::, IDNA liqase C!
A.-
S
eq
_ /7,
60
inated (^) with a small amount of RNase H and exonuclease, but this did not influence the results since the (^) nucleases were added in (^) excess. Also, ligation product observed in our reactions (^) did not contain any RNA nucleotide as determined by resistance to treatment with 1 M KOH following the joining reaction (data not shown). These reactions are a reconstitution of enzymatic activities required for the com- pletion of lagging-strand DNA replication. We have used DNA polymerase efor the complete reaction shown here. However, we emphasize that polymerase E is not (^) necessarily the (^) enzyme used (^) in vivo. The action ofRNase HI and 5'-to-3' exonuclease generates two DNA primers separated by a^ short gap hybridized to a DNA template. We have shown (22) that calf DNA polymerase a, 6/proliferating cell nuclease antigen (PCNA), or E is capable of closure of such a gap to a nick that can bejoined by DNA ligase I. Waga and Stillman (13) found that PCNA, replication factor C, and replication protein A blocked ligation in a substrate similar to that used here when DNA polymerase a was the only polymerase present. This indicated a need to switch to polymerase 8 or^ E. The mammalian 5'-to-3' (^) exonuclease, DNA (^) ligase I (12), and possibly RNase HI were found to (^) be necessary for formation of form^ I^ simian virus^40 DNA^ (11-13). Here we show the reactions by which RNase HI acts with the 5'-to-3'
exonuclease to remove the RNA primer ofOkazaki fragments.
In summary (Fig. 7), reconstitution of Okazaki-fragment
processing requires the action oftwo nucleases. The reaction
NCNCN.~~~~NC
4l---"~
B
L) rI~~ra se
,. (^) ><D
Ir,
XX>~~~~XX
4,
I: 3
FIG. 6. Complete lagging-strand processing by purified proteins. The (^) lagging-strand substrate was (^) prepared for this (^) experiment by hybridizing the 13-nucleotide^ RNA^ to^ the^ single-stranded 130- nucleotide template and extending and labeling the 3' terminus. The substrate was incubated with DNA polymerase e (0.1 units), DNA ligase I (0.1 units), RNase HI (0.3 units), and 5'-to-3' exonuclease (0.3 units) as shown in the figure. Reaction mixtures were incubated and processed as described in Fig. 2. The final ligation product, 122 nucleotides in length, is indicated by an arrow.
FIG. 7. Model for the completion of mammalian lagging-strand DNA (^) replication. The initial reaction involves a site-specific cleav- age by RNase^ HI^ at^ the^ phosphodiester bond of the^ nucleotide 5'^ of the RNA-DNA (^) junction. Removal of the last 5' ribonucleotide is performed by 5'-to-3' exonuclease and results in a 5' (^) phosphorylated DNA terminus. Action of the exonuclease is likely to be stimulated by the cleaved segment of RNA or by elongation of the upstream primer as shown. A nick is then generated between the two DNA termini, which are joined by DNA ligase I.
(^9806) Biochemistry: Turchi et al.
E
Proc. Natl. Acad. Sci. USA 91 (1994) 9807
is very specific in that on our substrate only two cleavages are required. RNase HI makes a specific endonucleolytic cut, leaving a single ribonucleotide that is removed by the 5'-to-3' exonuclease. The nucleases can act^ in the^ same^ reaction with DNA polymerase E and DNA ligase I to complete the reaction
of Okazaki-fragment processing. The simian virus^40 in vitro
DNA replication system has identified and assigned roles for a number of proteins involved in^ steps^ of^ eukaryotic DNA replication (1, 11-13). We have added to this knowledge by identifying steps involved in removal of initiator RNA prim- ers prior to joining of Okazaki fragments, representing the final steps in mammalian DNA replication.
This work was supported by National Institutes of Health Grant GM24441, by Cancer Center Core Grant CA11198, and^ by funds^ from Wright State University.
- Stillman, B. (1989) Annu. Rev. Cell Biol. 5, 197-245.
- Tsurimoto, T. & Stillman, B.^ (1991) J.^ Biol.^ Chem.^ 266, 1%1-1968.
- Eki, T., Matsumoto, T., Murakami, Y. & Hurwitz, J. (1992) J. Biol. Chem. 267, 7284-7294.
- Johnson, L. M., Snyder, M., Chang, L. M., Davis, R. W. & Campbell, J. L. (1985) Cell 43, 369-377.
- Boulet, A., Simon, M., Faye, G.,^ Bauer,^ G.^ A.^ &^ Burgers, P. M. (1989) EMBO J. 8, 1849-1854.
- Morrison, A., Araki, H., Clark, A. B., Hamatake, R. K. & Sugino, A.^ (1990) Cell^ 62, 1143-1151.
- Araki, H., Ropp, P. A., Johnson, A.^ L., Johnston, L.^ H., Morrison, A. & Sugino, A. (1992) EMBO J. 11, 733-740.
- Bambara, R. A. & Jessee, C. B.^ (1991) Biochim.^ Biophys. Acta 1088, 11-24.
- Burgers, P. M. (1991) J. Biol. Chem. 266, 22698-22706.
- Nethanel, T. & Kaufmann, G. (1990) J. Virol. 64, 5912-5918.
- Ishimi, Y., Claude, A., Bullock, P. & Hurwitz, J. (1988) J. Biol. Chem. 263, 19723-19733.
- Waga, S., Bauer, G. & Stillman, B. (1994) J. Biol. Chem. 269, 10923-10934.
- Waga, S. & Stillman, B. (1994) Nature (London) 369, 207-212.
- Kogama, T. (1986) J.^ Bacteriol. 166,^ 361-363.
- Goulian, M., Richards, S. H., Heard, C. J. & Bigsby, B. M. (1990) J. Biol. Chem. 265, 18461-18471.
- (^) Siegal, G., Turchi, J. J., Myers, T. W. & Bambara, R. A. (1992) Proc. Natl. Acad. Sci. USA 89,^ 9377-9381.
- Murante, R. S., Huang, L., Turchi, J. J. & Bambara, R. A. (1994) J. Biol. Chem. 269,^ 1191-11%.
- Lyamichev, V., Brow, M. A. D. & Dahlberg, J. E. (1993) Science 260, 778-783.
- Bfisen,^ W.^ (1980)^ J.^ Biol. Chem.^ 255,^ 9434-9443.
- Goulian, M. & Heard, C. J. (1991) Anal. Biochem. 192, 398-
- Bfisen, W., Peters, J. H., Hausen, P. & Crystalla, G. (1977) Eur. J. Biochem. 74, 203-208.
- Turchi, J. J. & Bambara, R. A. (1993) J. Biol. Chem. 268, 15136-15141.
- Eder, P. S. & Walder, J. A. (1991) J. Biol. Chem. 266, 6472-
- Grosse, F. & Krauss, G. (1985) J. Biol. Chem. 260, 1881-1888.
- Kornberg, A. & Baker, T. A. (1992) DNA Replication (Free- man, New (^) York).
- DiFrancesco, R.^ A.^ & Lehman,^ I.^ R. (1985) J. Biol.^ Chem.^ 260, 14764-14770.
Biochemistry: Turchi^ et^ al.